 'Livets tre' - felles avstamning
'Livets tre' - felles avstamning
(Oversatt fra 'Zombie Science' kap 3, av J. Wells)
Av alle villedende evolusjonsikoner, er livets tre det mest fundamentale. Vi vet intuitivt at noen dyr er mer lik andre utseendemessig, og folk har klassifisert organismer i århundrer, og resultatet er hierarkisk organisert: levende organismer, dyr, virveldyr, fugler etc. Taxonomi -navnsetting og klassifisering av dyr og planter, etter art og slekt ble grunnlagt av Carl Linné i det 18-århundre. Når en klassifiserer levende organismer, gjøres det etter likheter som kalles homologier. Noen ganger er disse likeverdige i struktur og posisjon, men kan utføre ulike funksjoner. I det 19-århundre atskilte Richard Owen (britisk biolog) homologier fra 'analogier', trekk som benytter ulike strukturer til å utføre lignende funksjoner. F.eks. inneholdt en fugls og en flaggermus sin vinge, ben av liknende posisjon og struktur, så deres vinger ble regnet som homologe. Et insekts vinge utfører samme funksjon, men har en annen struktur, så dens vinger regnes som analoge til dem hos fugler og flaggermus. (Bilde analoge strukturer)
Bilde 1. Skissert i 'Origin of Species'
Owen tilordnet homologi til konstruksjon av en felles plan, men Charles Darwin mente den beste forklaring til homologi var en nedstamning fra en felles stamfar. Darwin uttalte: "Således ser jeg alle vesener ikke som spesielle skapninger, men som lineære nedstigninger fra noen få organiske vesener som levde i fjern fortid." (2) Selv om Darwin var ambivalent m.h.t. hvorvidt det var separate stamfedre for hver stor klasse" eller en enkelt stamfar for alle levende vesener, så helte han mot sistnevnte alternativ. Han skrev at "sannsynligvis har alle organismer som noensinne har levd på jorda, nedstammet fra en tidligst begynnende form." (3) I Darwins tegnede livstre symboliserer de prikkede linjene Darwins ambivalens til hvorvidt det var én felles stamfar, noe han altså holdt på. (5)
For Darwin var evolusjon fullstendig materialistisk. Alle levende former etter den første var resultat av avstamning ved modifikasjon ved naturlig seleksjon. Og dermed skulle det ikke være noen diskontinuiteter i det påfølgende livets historie. I 1859 skrev han til geolog Charles Lyell, "jeg ville absolutt ikke gi noe for min teori om naturlig seleksjon, dersom den krevde mirakuløse tilføyelser på hvilket som helst nedstignings stadium." (6)
Vi skal se på noen grunner til at dette bildet ikke stemmer:
 Fossiler
Fossiler
Det "utenkelig store" antall av overgangs-linker som ble postulert av Darwin, har aldri blitt funnet. Darwin kjente til problemet i 1859, og han erkjente at det var et seriøst problem, som "visselig kunne erkjennes som et gyldig argument." (8) Han tilordnet problemet til ufullstendigheten av fossilmaterialet, men siden har problemet vokst. I 1991 konkluderte et team av paleontologer at den kambriske eksplosjon var endog mer plutselig og omfattende enn tidligere forespeilt." (9) En av forfatterne, James Valentine, som er evolusjonist, spesifiserer i forordet at "Darwin var korrekt i sine konklusjoner at alle levende organismer har nedstammet fra en felles stamfar, og kan bli plassert innenfor livets tre." (10) Men senere i boka, innrømmer Valentine at "organismer med den karakteristiske kroppsformen som vi identifiserer i dyrerekker, dukker plutselig opp i fossilregisteret, mange innen et kort vindu av geologisk tid .. Det er konsistent med fossilregisteret at alle karakteristiske dyrekropps-former hadde utviklet seg ved slutten av denne perioden, men ingen av dem kan spores gjennom fossile overgangsformer til en opphavelig gruppe." (11)
I 2013 publiserte Valentine og Douglas Erwin en annen bok om den kambriske eksplosjon. Erwin tror på evolusjonsteorien, liksom Valentine, og boka deres består av evolusjonære hypoteser vevd i en historie om hva de tror skjedde før og etter denne eksplosjonen. I prosessen benytter de 'spøkelses-linjer' -hypotetiske linker som ikke etterlater noe fossilspor, men hvis eksistens kreves ut fra forestillingen om felles avstamning. Erwin og Valtenine refererer endog ironisk til sin hypotese som "øvelse i evolusjonære seanser." (12)
Bilde 2. Fossil av Dourodon (
Men hva om de nye kroppsformene som oppsto i den kambriske eksplosjon -lenke, krevde ny informasjon, som er immateriell? I sin bok Darwins Doubt -her, argumenterer filosofiprosessor St. C. Meyer at den kambriske eksplosjon var en eksplosjon i ny informasjon. I følge dr. Meyer er den eneste kjente kilde til slik informasjon intelligens, så den kambriske eksplosjon er bevis for intelligent design. (13) Så bevisene kan si ja, men vitenskapen sier nei.
I 1972 kalte de evolusjonistiske paleontologene N. Eldridge og St. Jay Gould et slikt mønster for "punktert likevekt" (14) I følge Gould "visste hver paleontolog alltid" at dette er det dominerende mønsteret i fossilregisteret. (15) De to tilordnet likevekt til konstante omgivelser eller interne begrensninger i embryo-utviklingen, og de tilordnet plutselige tilsynekomster til 'allopatrisk' artsdannelse. (Selve ordet allopatrisk' kommer fra ord som betyr 'andre' og 'far'). I følge hypotesen om allopatrisk' artsdannelse' skal en liten del av en eksisterende populasjon blir geografisk atskilt, og slik atskilt fra hoved-populasjonen. Den lille atskilte grenen kan så via genetiske endringer (mutasjoner) vende det isolerte delen til en ny art. Men på grunn av liten størrelse og rask evolusjon, ville den ikke etterlate noe fossilregister. Dermed vil den virke som å oppstå plutselig.
Noen kritikere innvendte at dette bare var et forsøk på å bortforklare fraværet av bevis for overgangsformer. Faktisk mente noen kritikere at et mønster med punktert likevekt er mer konsistent med skapelse enn med evolusjon. Men Darwin hadde erklært denne ideen for uvitenskapelig. En annen innvending var at utvikling av slike morfologiske trekk, ved genetiske mutasjoner og naturlig seleksjon, ville kreve større populasjoner og lengre tidsperioder. Slike trekk kunne ikke forventes å oppstå i små populasjoner over korte tidsperioder {i hvert fall om ikke muligheter for endring lå innebakt i genomet -oversetters tilføyelse.} R. Lande og M. Slatkin skrev i 1982 at "noen av de genetiske mekanismene som har blitt foreslått for å forklare de plutselige oppdukkingene og forlengede perioder av 'stasis' (ikke-forandring), i mange fossilarter mangler kontinuerlig empirisk støtte." (16).
Hvorfor fossiler ikke kan etablere stamfar-avstamning relasjoner
I følge den britiske biologen Ronald Jenner, så er det "få andre valg enn å omsortere ut fra våre mer eller mindre informerte forestillingsevne, for å produsere den historiske beretningen som er det endelige målet for våre studier om dyrs utvikling." Faktisk er "vår forestillingsevne det eneste verktøy som kan flette fragmentariske bevis til en sømløs historisk beretning, som relaterer 'hva, hvordan og hvorfor' i forbindelse med evolusjon." (17) Situasjonen er faktisk verre enn det. Selv om vi hadde gode fossil-registre, ville vi likevel trenge å bruke vår forestillingsevne til å produsere beretninger om 'stamfar-avstamning' relasjoner.
Om en fant to menneskelige skjelett begravd i en åker, hvordan kunne en vite hvorvidt det ene var i slekt med det andre? Uten identifikatorer, noe skriftlig eller i noen tilfelle DNA, ville det være umulig å vite. Likevel ville det dreie seg om to skjelett fra det samme nålevende art. Med to ulike, gamle, utdødde arter, ofte fjernet fra hverandre i tid og rom, så ville det ikke være noen måte å demonstrere en foreldre-avkom relasjon.
For flere tiår siden skrev paleontolog ved American Museum of Natural History, Gareth Nelson, "Forestillingen om at man kan gå til fossil-registeret og forvente empirisk å opprette en sekvens av stamfar-etterkommere .. har vært og fortsetter å være en skadelig (pernicious) illusjon." (18) I 1999 skrev redaktør i Nature, Henre Gee at: "det er faktisk i realiteten umulig å linke fossiler til kjeder og virkning på noen gyldig måte." Han konkluderte: "Å ta en serie fossiler og hevde at de representerer en linje, er ikke en testbar vitenskapelig hypotese, men en forutsetning som bære samme gyldighet som en 'godnatt-fortelling' -morsom, kanskje til og med lærerik, men ikke vitenskapelig." (19) Gee tror (i likhet med Valentine og Erwin) på evolusjonsteorien, men hans tro er i likhet med deres ikke basert på fossilbevis.
Fylogenetiske trær
Til tross for at avstamnings-relasjoner ikke kan bli empirisk gjenvunnet ved fossiler, er den moderne biologiske litteraturen full av evolusjonære trær, kalt 'fylogenetiske trær', som formodentlig skal vise nære relasjoner. Trærne blir typisk ledsaget med fortellinger om hvordan tidligere organismer utviklet seg til senere. Men fylogenetiske trær krever ikke stamfedre. Faktisk så krever de ikke engang organismer.
I 2013 produserte en vitenskapelig utdanningsgruppe en leksjonsplan for å undervise videregående skoler og høyskole -studenter hvordan en skulle konstruere et fylogenetisk tre .. Selv om objektene der var kunstige, så var "problemet som ble frontet og spørsmålene stilt, lik dem som adresseres av paleontologer som benytter fossil-mønstre." (20) Det første prinsippet å styre etter er: "Organismer som likner hverandre på mange måter, er sannsynlig mer relaterte enn organismer som bare ligner hverandre litt. Det vil si: Dess større likhet i struktur (dess flere felles trekk), dess nærmere sannsynlig slektskap (relationship) mellom de to formene." Studenter anbefales å "ta den minste, enkleste form som den sannsynlige felles stamfar for gruppa og så forsøke å arrangere de andre som grener på ett tre ut fra den stamfaren." (21)
Fylogenetiske trær benyttes mer for å konstruere 'nærhets-forbindelser', som at 'jern står nærmere aluminium, enn de begge er i forhold til ei påskelilje. Men mange evolusjonister blir utydelige i å foreslå at de har demonstrert slektskap i genetisk henseende.
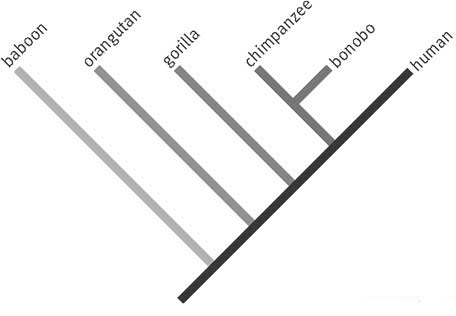 Kladogram
Kladogram
I 1950 grunnla den tyske biologen Willi Hennig feltet fylogenetisk systematikk, senere kalt kladistikk. ('Clade kommer fra et gresk ord som betyr 'gren'.) Hennig trodde at evolusjonsteorien var et faktum, men han mislikte {i likhet med oss -oversetters kommentar} ubegrenset evolusjonær historiefortelling, og han søkte et mer rigorøst empirisk måte å etablere relasjoner mellom levende organismer. (22) I kladistikk, er et 'kladogram' et diagram laget ved å sammenligne karakteristika delt av visse grupper av taxa (organismer, slik som arter, slekter, familier etc); men ikke av en annen taksonomisk gruppe. Førstnevnte kalles 'inngruppen', den andre kalles 'utgruppen'. Karakteristika funnet i inngruppen, men ikke i utgruppen sies å være avgrenet (derived). Resultatet blir et nøstet hierarki. Kladistikere regner denne tilnærmingen som mer presis og testbar enn ganske enkelt å telle likheter.
Det er verken tidsdimensjon eller slekts-relasjoner i et kladogram. En evolusjonsbiolog kan forsikre at utgruppen og inngruppen er beslektet gjennom stamfar og avstamning, men det er ikke noe i et kladogram i seg selv som krever en slik relasjon. Det gammeldagse evolusjonære treet som ble benyttet til å pryde og smykke lærebøker i biologi, har nå i hovedsak blitt erstattet av kladogram. Likevel villeder enda mange lærebøker studenter til å tro at kladogram er stamfar-avhengige diagram. F.eks. i K.Miller og J. Levines mye brukte 'Biology', hevder at kladogram linker grupper av organismer ved å vise hvordan evolusjonære avstamningslinjer delte seg ut fra felles stamfedre." (24) Men de felles stamfedrene ser en ikke noe til.
Molekylær fylogeni
Siden opprinnelsen av molekylær biologi i midten av det 20. århundre, har biologer i økende grad benyttet sammenlikning av sekvenser i DNA, RNA og proteiner til å konstruere fylogenetiske trær. F.eks. en spesiell DNA-sekvens kunne være tilstede i ulike arter, med mindre variasjoner. Å sammenlikne sekvens-ulikheter i arter A, B og C kan føre til at en finner at A er mer lik art B enn art C. Likheten mellom de to sekvensene, ofte kalt homologi, kan uttrykkes som en prosentdel, som representerer hvor mange underenheter på tilsvarende posisjoner, som er identiske mellom dem.
Likhet kan antas å implisere slektskap, men det er bare en formodning. Enhver slutning til avstamning basert på sekvens-likhet er hypotetisk. Og siden molekylære sekvenser, med sjeldne unntak er tilgjengelige bare fra levende organismer, så blir enhver slutning om den evolusjonære fortiden til disse organismene, inkludert deres stamfedre, enda mer hypotetisk.
Sammenstillings (alignment) -problemet
Før sekvenser kan sammenlignes, må de sammenstilles (be aligned). Men molekylære sekvenser i levende organismer inneholder typisk repeterende segmenter. Det innebærer at det ofte er uklart hvor en skal starte å sammenligne dem. Segmenter kan være ganske lange, og siden de kan sammenstilles på mer enn en måte, avhenger resultatet tungt av hvilken sammenstilling undersøkeren benytter. Og når det er mange sekvenser som sammenlignes, som det er i molekylær fylogeni, så blir problemet verre. (25)
Dataprogram er tilgjengelige til å sammenstille sekvenser, men de avhenger av parametrene som blir innsatt av programmererne, og noen ganger er resultatene biologisk usannsynlige. I 2009 undersøkte biolog David Morrison den vitenskapelige litteraturen og fant at "mer enn halvparten av evolusjonære biologer griper manuelt inn i sin sekvens-sammenstilling, og flere enn 3/4 av fylogenetikere gjør det." (26) I 2015 la Morrison merke til "en knoppskyting av sammenstillings metoder" som "produserer oppdagbare ulike multiple sekvens-sammenstillinger i nesten alle realistiske tilfeller." (27) Så selv før selve prosessen ved å sammenlikne sekvenser starter, influerer subjektive valg de eventuelle resultatene. Likheter forutsettes å fortelle undersøkerne hvem som kan relateres til hvem og hvor nært. Men slik det viser seg, gjør undersøkerne mye av varslingen ved å velge og vrake sine foretrukne sammenstillinger.
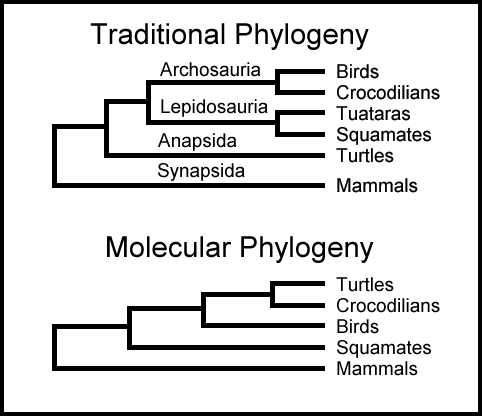 Trær i Konflikt
Trær i Konflikt
Det er ikke rart da at sekvens-sammenstillinger ofte produserer fylogenetiske trær i konflikt med hverandre. For noen tiår siden var det antatt av inkludering av flere molekyler i analysene ville eliminere ulikhetene, men det har ikke skjedd. I 2005 analyserte evolusjons-biologene A. Rokas, D. Krüger og Sean B. Carrol femti gener fra 17 dyregrupper og konkluderte at "ulike fylogenetiske analyser kan nå motsigende resultater med [tilsynelatende] absolutt støtte." (28)
I 2008 benyttet et internasjonalt team på 18 biologer 150 gener til å konstruere en fylogeni av dyregrupper. Deres tre ble motsagt følgende år av et annet internasjonalt team av 20 biologer som benyttet 128 gener. (29,30) I 2012 rapporterte biologene L. Dávalos, A. Cirranello, J. Geilser og N. Simmons: "Inkongruens mellom fylogenier oppnådd fra morfologiske vs. molekylære analyser, og mellom trær basert på ulike sub-sett av molekylære sekvenser har blitt gjennomgående." (31)
Bilde 5. Fort uoverensstemmelser i ulike fylogenier
For å eliminere konflikter mellom molekylære fylogenier, ekskluderer biologer ofte data fra sine analyser, og rettferdiggjør praksisen på grunn av at ikke alle sekvenser bærer "sterke fylogenetiske signaler." (32) F.eks. taksonomiske grupper sluttes vanligvis å være nært relatert, dersom deres korresponderende sekvenser er lignende, men sekvenser kan være meget ulike selv i taksonomiske grupper som er antatt av andre grunner nylig å ha skilt seg ut fra en felles stamfar. De avvikende sekvensene antas da å være "raskt utviklende" og blir ekskludert fra det fylogenetiske treet. (33) Eller korresponderende sekvenser kan være meget like i taksonomiske grupper som antas å ha vært lenge atskilt, i hvis tilfelle deres likhet tilordnes å ha akkumulert tilstrekkelig antall mutasjoner til å konvergere mot hverandre (et fenomen som kalles "lang-gren tiltrekning") (34)
Bemerk hvor mye formodningene spiller inn i disse argumentene. Alle molekylære sekvenser (med noen få unntak) er fra levende organismer. Det er ingen måte å se dypt ned i fortida og bestemmer hvorvidt de avvek raskt eller konvergerte sakte. Så en fylogenetiker formoder at evolusjonsteorien er sann, og at livets historie er tre-liknende. Så velges et spesielt fylogenetisk tre og forkaster de sekvenser om ikke passer inn. (Fig.2-4). Det blir 'felles avstamning, unntatt når det ikke er det.'
 Orfan-gener
Orfan-gener
Praksisen med å forkaste ikke-ønskede sekvenser er blitt mer fremtredende på grunn av nylige oppdagelser at mange DNA-sekvenser er begrenset til en enkelt taksonomisk gruppe.
I følge evolusjonsteorien stammer alle nye sekvenser fra gamle ved duplisering og/eller mutasjoner samt naturlig seleksjon. I 1977 skrev den franske molekylær-biologen Fr. Jacob: "Da livet hadde startet i form av noen primitive selv-reproduserende organismer, så måtte videre evolusjon fortsette hovedsakelig gjennom endringer av allerede eksisterende sammensetninger." Derfor: "Dannelsen av helt nye DNA-sekvenser kunne ikke være av noen betydning i produksjonen av ny informasjon" (35) Så en sekvens i art X skulle ligne dens opprinnelige sekvens i art Y.
I løpet av 1990-årene imidlertid hadde biologer oppdaget mange sekvenser med ingen likhet til dem i andre taksonomiske grupper. I 1999 undersøkte D. Fischer og D. Eisenberg den komplette DNA-sekvensen til mer enn et dusin arter av bakterier og konkluderte med at ca. 1/3 av protein-kodende regioner ikke hadde noen likhet til proteiner fra andre genom. (Genom - som sammenstillingen av hele organismens DNA). Dette stiller et ukomfortabelt spørsmål til evolusjonsteorien. Fischer og Eisenbert spurte: "Dersom proteiner i ulike organismer har utviklet seg fra felles opprinnelige proteiner ved duplisering og tilpasset variasjon, hvorfor viser så mange i dag ingen likhet til hverandre?" Godt spørsmål.
Bilde 6. Problemstilling
Siden protein-kodende regioner også er kjent som "Open Reading FrAmes", kalte . Fischer og Eisenberg disse for 'ORFAns" (36) Antallet av kjente ORFAns har vokst helt siden da. De er blitt oppdaget i gjær, bananfluer (40), mus (41) og mennesker (43, 44) En artikkel fra 2015 i Nature rapporterte hundrevis av ORFans begrenset til blekksprut. (45) Faktisk har hver art, hvis genom er undersøkt, hatt et signifikant antall ORFans. Biologer venter nå å finne ORFans hvor de enn leter. Hvor kommer ORFans fra? Den tyske biologen H. Kaess-mann skrev i 2010 at et gen som synes å dannes fra 'scratch' trolig oppstår gjennom en "tidligere ikke-funksjonell genomisk struktur", ikke relatert til noe pre-eksisterende genetisk materiale." (46) I 2011 argumenterte biologene D. Tautz og T. Domazet-Loso på ligende vis at "mange ORFAns kan ha oppstått 'de novo' fra ikke-kodende regioner." (47) Men "kan ha oppstått de novo". Det er ikke så mye bedre enn å si at ORFAns 'bare dukker opp'.
Oppmerksomme på problemet foreslo en internasjonal gruppe i 2012 en evolusjonær modell, der funksjonelle gener utvikles de novo gjennom forbigående proto-gener." (48) Liksom den evolusjonære historien krever "spøkelses-linjer" til å stoppe gapet mellom fossiler, så krever den innbilte "proto-gener" til å bygge gapet mellom ORFans og ikke-kodende regioner. {I tilfelle ville muligheten måtte være lagt inn fra starten av -intelligent formet.}
Så det er vanskelig å forklare opprinnelsen til ORFans. Det er enda vanskeligere å ha med dem å gjøre når en skal konstruere fylogenetiske trær. Forfatterne til en artikkel i 2015 om insekt-fylogeni, besluttet å behandle dem ved å ignorere dem fullstendig. De forklarte at "vår tilnærming eliminerer ethvert gen som er nærværende i bare en taksonomisk gruppe" fordi "slike gener er fylogenetisk ikke-informative". "Ikke-informative" det er hvis den aktuelle arten er forutsatt å ha nedkommet fra en felles stamfar. Kanskje "anti-informativ" ville være et bedre ord i den sammenheng. Så forfatterne plukket ut godbitene fra hvert insekt de studerte, og kastet 40% av sekvensene fra frukt-flua og 80% av sekvensene fra vannloppa for å danne et fylogenetisk tre de kunne like. (49)
Metoden med å konstruere fylogenetiske trær, kan bli oppsummert slik: 1. Anta at felles avstamning er sant. 2. Plukk ut godbitene av data som inkluderer ulike organismer 3) Konkluder at de organismene er i slekt med en felles stamfar. Det er ikke empirisk vitenskap. Det er mer som å spå i te-blader.
Tilfellet med den manglende stammen
Bilde 7. 'Livets tre' kan falle tilbake på neo-darwinismen
Om det fantes ett enkelt livets tre, så ville stammen ha vært en enkelt "universell stamfar". Men ulikheter i molekylær fylogeni har overbevist neon vitenskapsfolk at livets historie ikke kan bli representert ved ett enkelt tre. Bevisene for en felles stamfar mangler. Og noen evolusjonister har avgjort at bevisene mangler, fordi stammen på treet er en luftspeiling.
Levende celler var tradisjonelt kalssifisert i to brede domener, prokaryoter (uten membranbundet kjene) og eukaryoter (med kjerne). De første gjaldt bakterier og den siste inkluderte en-cellede organismer som gjær, likeså flercellede organismer som planter og dyr. Basert på molekylær sammenligning imidlertid, rapporterte Carl Wose og George Fox i 1977 at noen prokaryoter er så radikalt ulik bakterer at de burde klassifiseres i et 3.domene, som de kalte arke-bakterier (archaea)." (50) Woese konkluderte med at forskjellene mellom archaea og bakterier og mellom dem og eukaryoter var for stort til å bli forklart med avstamning fra en felles stamfar, slik termen vanligvis forstås. "Stamfaren kan ikke ha vært en spesiell enkelt-cellet organisme, i en enkel organisatorisk linje," skrev Woese i 1998. I følge Woese var de primitive cellene, som han kalte progenotes, meget ulike moderne celler. En 'progenote' var mer lik en sekk med semi-autonom genetiske elementer" som "ville komme og gå", så progenoter var ikke organismer i noen konvensjonell betydning av ordet. (52)
Men om vår felles stamfar med archaea og bakterier ikke var en organisme, på hvilken måte var den en stamfar. Hvorfor stoppe med hypotetiske progenoter? Hvorfor ikke si at vår felles stamfar var ursuppen? eller elementene i den periodiske tabellen?
Likevel fortsatte evolusjonsbiologer, selv de som delte Woeses syn, å forsvare ideen om en felles stamfar. F.eks. skrev W.F. Doolittle i 2009 at han betvilte at det noensinne var en enkelt universell felles stamfar." men "dette betyr ikke at livet mangler 'universell stamfar-skap', fordi 'stamfar-skap' inneholder ikke en felles stamfar. Hvorfor slik mental gymnastikk? Doolittle innrømmer åpent at det er på grunn av at 'det står mye på spill sosiopolitisk', nemlig behovet for å slå 'anti-evolusjonistene' i 'den kulturelle krigen'. (53)
Så vi kan ikke slutte til stamfedre fra fossiler og det er ingen i kladogram. Molekylære fylogenier, selv med alle sine metodologiske problemer og konfliktfylte resultater, kan bare foreslå hypotetiske stamfedre. Og et sammensurium av en 'ikke-organisme' er ikke en stamfar i noen meningsfull betydning av ordet. Alle stamfedrene mangler, men zombie-science forsetter å insistere på at de må ha eksistert.
 Homologi
Homologi
Vi har tidligere vært inne på dette, og kom til at en måte å definere homologi (samme-si) er å definere homologi som likhet i struktur eller posisjon. Darwin så homologi som bevis for sin teori, i det han listet det i 'The Origins of Species' blant de fakta som proklamerte så klart at "proklamerer så klart at de utallige arter, slekter og familier med organiske vesener, som denne verden er full av, har alle nedkommet, hver innen sin egen klasse eller gruppe, fra felles foreldre." (54) Men hvordan ble homologe trekke overført fra foreldre til etterkommere? Benene i en fugls ving er ikke tilstede {som sådanne -oversetters kommentar} i egget. I følge en hypotese er mekanismen genetisk: Strukturene er homologe fordi de er spesifisert ved lignende gener. Bevis støtter ikke denne hypotesen, imidlertid, og biologer har visst det i flere tiår. I 1971 skrev den britiske embryologen Gavin de Beer: "Karaktertrekk kontrollert av identiske gener er ikke nødvendigvis homologe" og "homologe strukturer trenger ikke kontrolleres ved identiske gener." (55)
I følge en annen hypotese er mekanismen en utviklende en: Strukturer er homologe fordi de oppstår fra lignende celler i embryoet. Men igjen støtter ikke bevis hypotesen. Så langt tilbake som 1958 , bemerket de Beer: "Korresponderende homologe strukturer kan ikke bli ført tilbake til lignende posisjon av cellene i embryoet, eller til deler av egget der strukturene blir satt sammen eller fra utviklingsmekanismer som de blir dannet fra." (56)
Homologi som felles avstamning
I fravær av en biologisk mekanisme til å danne homologi, redefinerte darwinister homologi til å bety likhet grunnet felles avstamning. I 1982 skrev Harvard biolog Ernst Mayr at det er bare en definisjon av homologi som gir biologisk mening: "Egenskaper til to organismer er homologe når de er oppnådd fra et lignende karakteristika av den felles stamfaren." (57) Således er i følge Berkeley evolusjonsbiolog D. Wake: "felles avstamning alt som definerer homologi." (58) Men å definere homologi som felles avstamning, fører til et seriøst problem: Med en gang homologi defineres som felles avstamning, kan det ikke lenger bli benyttet som bevis for felles avstamning. Det ville innebære å resonere i sirkel: Hvordan vet vi at trekk B stammer fra trekk A? Fordi trekk B er homologt med A. Hvordan vet vi at det er homologt med A? Fordi B nedstammet fra A. Så for å få det en ønsker, må en ha det allerede.
Dette logiske problemet med homologi (definert som likhet grunnet felles avstamning) har vært kjent i flere tiår, men de fleste nåværende lærebøker ignorerer det. Miller og Levine's lærebok 'Biology' fra 2014 definerer homologi som trekk som "har blitt arvet fra en felles stamfar" og to sider senere står det: " homologe molekyler gir bevis for felles avstamning." (59) I følge S. Mader og M. Windelspecht's 2016 lærebok 'Biology': "Strukturer som er anatomisk like fordi de arvet fra en felles stamfar, kalles homologe. I kontrast er analoge strukturer som tjener samme funksjon, men oppsto uavhengig i ulike organisme-grupper som ikke deler en felles stamfar. Vingene til fugler og insekter er analoge strukturer. Således er homologe , ikke analoge, strukturer bevis for felles avstamning for spesielle grupper av organismer. " (60) Det er igjen sirkulært resonnering maskert som bevis.
Bilde 8. Forvirring ang. analogi = homologi?
Et nåværende forsøk på å løse forvirringen
I 2014, publiserte Yale biolog G. Wagner en bok kalt 'Homology, Genes and Evolutionary Innovation'. I følge Wagner vil "enhver biolog være enig i at homologi er et forvirrende emne. " og han skrev boken for å prøve å redusere forvirringen. (61) Hans forutsetning i oppstarten var at "homologi er en hypotese om nedstamning fra en felles stamfar." og han bekreftet de Beer's poeng at homologi "ikke kan bli forklart ved identiske sett gener som styrer deres utvikling." (62) Likevel er det gener og ikke morfologiske trekk som arves fra en generasjon til neste. Så Wagner tilordnet homologi til utviklings gen-regulerende nettverk som "lå under evolusjonen av utviklingsveier, og således utvikling av morfologiske strukturer." (63) Så homologi har en genetisk basis når alt kommer til alt. (64)
Wagners oppstart-forutsetning førte ham til en annen paradoksal konklusjon: Homologi betyr ikke lenger likhet. Som vi så ovenfor, kan korresponderende DNA sekvenser være meget ulike i to arter, som antas å ha en nylig felles stamfar. Om homologi defineres i termer av felles avstamning, så er etterkommernes sekvenser homologe selv om de er ulike. Det samme kan skje ved morfologiske trekk. "Identiteten til et morfologisk karaktertrekk er ikke knyttet til likhet," skrev Wagner, men heller til "historisk kontinuitet for avstamning." (65) Likevel uten å benytte likhetstrekk, hvordan kan evolusjonsbiologer konstruere fylogenetiske trær? Og uten fylogenetiske trær, hvordan kan de slutte kontinuerlig avstamning?
Wagners bok var et ærlig forsøk på å rette opp i forvirringen omkring homologi. Men når homologi en gang er definert i termer av felles avstamning, synes det som forvirringen er uunngåelig.
Biolog-filosof R. Brady skrev for tre tiår siden: "ved å gjøre vår forklaring (felles avstamning) til definisjon av forholdene som skal forklares (homologi), uttrykker vi ikke vitenskapelige hypoteser, men tro. Vi er så overbevist at vår forklaring er sann at vi ikke lenger ser behov for å skjelne det fra situasjonen vi forsøker å forklare. Dogmatiske forsøk av dette slag må omsider forlate (empirisk) vitenskaps sfære." (66)
Bilde 9. Forsøk på å forklare 'konvergent evolusjon'
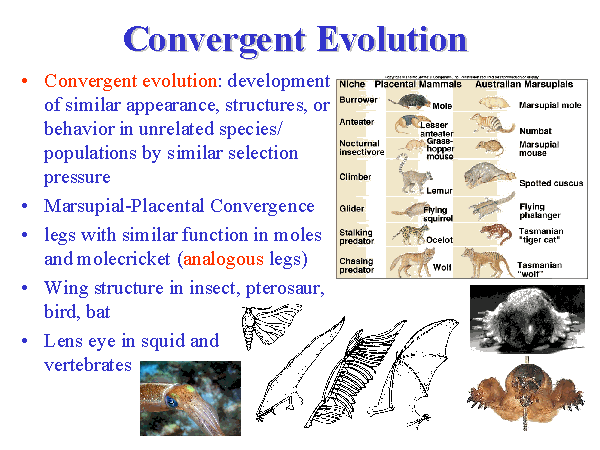 Konvergens
Konvergens
Eksistensen av mange likheter som helt klart ikke er resultat av felles avstamning. Et klassisk eks. er den slående likheten mellom kamera-øyet til et virveldyr og kamera-øyet til en blekksprut, som ingen tror er arvet fra en felles stamfar. Slike merkelige likheter var kjent i Darwins tid, og evolusjonsbiolog Ray Lankester foreslo i 1870 å erstatte 'homologi' med to nye termer: 'homogeny'-som betød likhet som skyldtes felles stamfar og 'homplasy' som betød likhet som ikke skyldtes felles avstamning. (67) Men 'homogeny' slo ikke an, og biologer fortsatte å benytte homology. Men homoplasy brukes nå {av noen- oversetters innskudd} for å referere likhet som ikke skyldes felles avstamning. Tydeligvis er likhet bevis på felles avstamning, unntatt når det ikke er det.
En annen term som ofte brukes i stedet for 'homoplasy' er 'konvergens'. To nylige bøker har listet hundrevis av eks. på konvergens i et vidt omfang av organismer: paleontolog S. Conway Morris (Life's Solution -2003) og morfolog G. McGhee's 'Convergent Evolution' (2011). Eksemplene på konvergens er mange. Conway Morris går så langt som til å si at "konvergens er meget vanlig." (ubiquitous) (68)
Dyrs konvergens
Pattedyr har tre måter de føder på: placenta-pattedyr nærer et foster via en morkake og føder fullt utviklede barn. Pungdyr gir fødsel til fostre som klatrer inn i en pung, der de fullfører fosterutviklingen. Og kloakkdyr legger egg i stedet for å føde levende fostre/barn. Fordi måtene som fødsel skjer på er så ulike, tror evolusjonærbiologer at disse tre gruppene atskilte seg lenge før de fleste av deres moderne karakteristika oppsto. Likevel fremviser pattedyr i disse tre gruppene utbredt konvergens: kamel-kenguru (flere mager, drøvtygging); ryggfinner hos haier og delfiner-se flere eks. bilde 9.
Plante-konvergens
Konvergens er vidt utbredt i planter også. Flere eks. opptrer i kjøttetende planter. 'Fallgruve' planter har et hulrom med nektar dypt inni seg, og glatte vegger. Insekter som tiltrekkes av nektaren, faller i fella og kan ikke komme ut. 'Fluepapir' planter har blader som skiller ut en tykk, lim-lignende substans som gjør insekter ubevegelige. I begge tilfeller blir så insektet fortært av plante enzymer eller mikroorganismer, og plantene absorberer næringsstoffene. 'Fallgruve' planter dukket tydeligvis opp seks ulike ganger, og 'Fluepapir' planter fem ulike ganger. (69) Verken 'Fallgruve'- eller 'Fluepapir' -planter nedstammet fra felles stamfar, som besatt deres distinktive kjøttetende trekk.
Et spesielt slående eks. på konvergens involverer planter i kaktus-familien i Amerika og vortemelk-familien i Afrika. Slike planter har tykke, 'kjøttfulle' stenger til å lagre vann og nåler i stedet for blader, men de oppsto tilsynelatende hver for seg. Kaktus og vortemelk finnes i tørre områder, som tilsynelatende er liknende, og evolusjonær teori tilegner deres konvergens til lignende omgivelser. Likevel påpekte et internasjonalt team i 2013 på at tidligere studier "inkluderte bare kvalitative beskrivelser av omgivelsene eller dårlige klimatiske målinger," men "de har latt utestet den avgjørende forutsetning om liknende omgivelses-press." Følgelig gjorde teamet et detaljert studium over omgivelsene til kaktus og vortemelk. Deres studium viste at planter "opptrådte i områder med ulike klimaer", som "skilte seg i større detaljer med sin nedbør og temperatur regimer." Faktisk var "likhetspunkter færre enn ulikhetene." (70) Hvorfor konvergerte plantene da til å bli så like?
Hva forårsaker konvergens?
G. McGhee argumenterer for at: "synet at evolusjon kun er historisk betinget, og således uforutsigbart (og ikke-repeterende), er demonstrativt falskt." Han konkluderer: "Fenomenet konvergent evolusjon demonstrerer at livet gjentatte ganger utvikler seg i et endelig antall av foretrukne retninger." (71) Om dette er sant, og om konvergens ikke skyldes naturlig seleksjon i lignende klima, hva skyldes det?
For S. Conway Morris skyldes konvergens biologiske begrensninger som leder embryo-utvikling i spesifiserte retninger (pathways). "Livets begrensninger gjør oppdukkingen av ulike biologiske egenskaper meget sannsynlig,"(72) argumenterer han. Således er utbredt konvergens å forvente. Så går han et steg videre: "Ikke bare er universet merkelig beredt for hensikt," skriver han, "men, også .. er livets evne til å navigere til sine løsninger." (73) Hensikt? Høres mistenkelig likt som design.
 Ett begrep maskert som et faktum
Ett begrep maskert som et faktum
Ironisk nok, ettersom evolusjonsbiologi har blitt mer empirisk som vitenskap, så har livets tre blitt mer illusorisk. Likevel i hovedstrøms biologi-lærebøker, og i populære TV-show og vitenskapelige magasiner, presenteres livets tre som et ubestridelig faktum. Hvorfor?
En grunn er at, så langt vi kan si, alle levende organismer stammer fra andre levende (likedannede) organismer. Plutselig nydannelse har aldri blitt observert, og vi ser avstamning bare innen eksisterende arter. Siden alle levende organismer kom fra andre levende organismer, er det ikke ufornuftig å tenke at kanskje noen arter kan ha nedstammet fra andre arter. Men hvorfor alle?
Grunnen til at vitenskap sier at alle arter må ha nedstammet fra felles stamfedre, er at naturalistisk vitenskap hater ideen at noen av dem ble skapt. I følge materialistisk vitenskap er skapelse utelukket fra starten av. Dersom (som Darwin tenkte) at eneste alternativ til livets tre, er separate skapelses-handlinger, og dersom skapelse ikke er tillatt, så vinner livets tre, uansett bevis.
Legg merke til at slik argumentering ikke kan forsvares på grunnlag av metodologisk naturalisme. Noen som opererer innenfor rammeverket til metodologisk naturalisme kunne beslutte at livets sanne historie {spesielt opprinnelsen -oversetters tilføyelse} er utenfor rekkevidde for empirisk vitenskap, og bevege seg over til mer sporbare fenomener. Ingenting tvinger slik en person til å anta en materialistisk konklusjon som har blitt avvist av tilgjengelig evidens. Evolusjonister som hevder at det 'må' eksistere et livets tre, uansett fakta: fossiler kan ikke produsere evidens for avstamning, uavhengig av konsistente motsetninger i molekylære fylogenier, og uavhengig av logisk bevis mot materialistiske forklaringer på homologi. De som tviholder på et slikt tre gjør det, ikke ut fra metodologisk naturalisme, men ut fra filosofisk naturalisme. Og verre, de stiller det fram som empirisk bevis, når det i virkeligheten er 'zombie-science'.
Livets tre er illustrasjon på en idé tenkt i materialistisk vitenskap, men har blitt mer enn det. Det er et ikon som har blitt brukt, og fortsetter å bli brukt, for å indoktrinere mennesker i det evolusjonære dogmet. Selv om St. Jay Gould trodde på evolusjon, skrev han innsiktsfullt i 1989: "Overtalelse ut fra ikoner, virker enda sterkere enn ord på kjernen i vårt vesen .. Men mange av våre bilder er inkarnasjoner av begreper som maskerer seg som nøytrale beskrivelser av naturen. De er de mest kraftfulle kilder til konformitet." (74)
Utvalgte Referanser:
26. Br. Alberts, “Science and human needs,” 137th Annual Meeting of the National Academy of Sciences, Washington, DC, May 1, 2000. Lenke.
27. Donald D. Hoffman og Chetan Prakash, “Objects of Consciousness,” Frontiers in Psychology 5 (2014):577.doi:10.3389/fpsyg.2014.00577.PMID:24987382.
Stoffutvalg og bilder ved Asbjørn E. Lund